IGEM:Imperial/2010/Protein Display
<html>
<body style="background-color:FFFFCC">
Receptor and Surface protein model
The aim of this model is to determine the concentration of Schistosoma elastase or TEV protease that should be added to the bacteria to trigger the response. It is also attempted to determine how long it will take for the protease or elastase to cleave enough peptides.
Cleavage of protein is an enzymatic reaction, which can be written as:
- E+S ↔ ES → E+P
- Substrate (S) = Protein
- Enzyme (E) = TEV (Protease)
- Product (P) = Peptide
This can be modelled in a very similar way to the 1-step amplification model, however, all the constants and initial concentrations will be different.
[TEV](t=0) - initial concentration of TEV will be arbitrarily chosen. However, ultimately we would need to measure the concentration of elastase that schistosoma releases.
Threshold concentration of peptide (20/08/2010)
The optimal peptide concentration required to activate ComD is 10 ng/ml <a href="http://ukpmc.ac.uk/backend/ptpmcrender.cgi?accid=PMC40587&blobtype=pdf">[1]</a>. This is the threshold value for ComD activation. However, the minimum concntration of peptide to give a detectable activation is 0.5ng/ml.
We want to know how long it takes until the threshold is reached:
- The mass of a peptide is 2.24kDa = 3.7184x10-21g.
- The number of molecules in one ml is 10ng/3.7184x10-21g = 2.6893x1012. In a litre, the number of molecules is 2.6893x1015.
- Dividing this value by Avogadro's constant gives the threshold concentration of cth=4.4658x10^-9 mol/L.
- The threshold for minimal activation of receptor is 2.2329x10-10 mol/L.
Protein production in B.sub (23/08/2010)
- The paper mentions that each cell displays 2.4x105 peptides. <a href="http://onlinelibrary.wiley.com/doi/10.1111/j.1574-6968.2000.tb09188.x/pdf">[2]</a>
- 2.4x105 molecules = 2.4x105/6.02x1023 mol = 0.398671x10-18 mol
- Volume of B.sub: 2.79x10-15 dm3</math>
- Concentration = [mol/L]
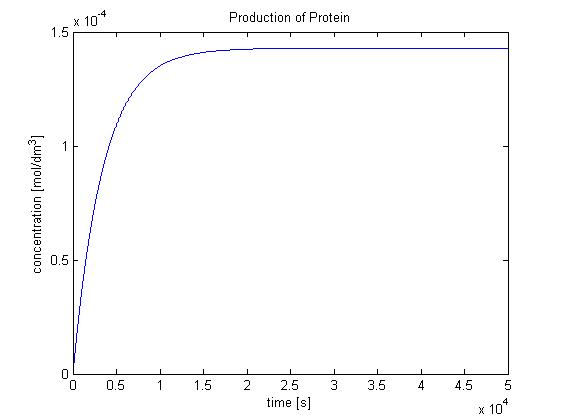
- c = 1.4289x10-4 mol/dm3. This is the concentration of protein that will be produced in a single cell of B.sub.
Hence, we can deduce that the concentration that the protein expression will tend to: c = 1.4289x10-4 mol/dm3 = cfinal.
Therefore, we can model the protein production by transcription and translation and adjust the production constant so the concentration value will tend towards cfinal.
Using a similar model to the simple production of Dioxygenase for the Output Amplification Model (Model preA), we obtain the following graph:
<img src="http://www.openwetware.org/images/f/f5/Protein_production.jpg" width="500" alt="Production of protein by transcription and translation"/>
The degradation rate was kept constant, and the production rate was changed according to the final concentration.
Control volume initial choice (23/08/2010)
All enzymatic reactions that we have modelled so far were confined whithin the bacterial cell. However, this case is different because the molecules are not confined by the bacterial membrane and can diffuse out of the cell.
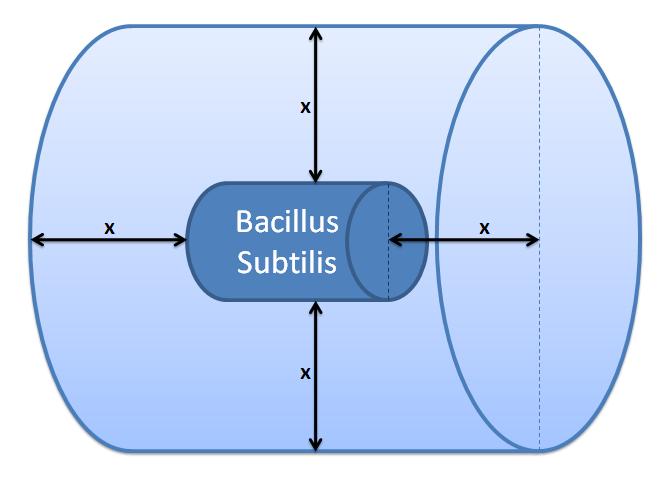
The control volume:
The inner boundary is determined by the bacterial cell (proteins after being displayed and cleaved cannot diffuse back into bacterium). The outer boundary is more time scale dependent. We have assumed that after mass cleavage of the display-proteins by TEV, many of these AIPs will bind to the receptors quite quickly (eg. 8 seconds). Our volume is determined by the distance that AIPs could travel outwards by diffusion within that short time. In this way, we are sure that the concentration of AIPs outside our control volume after a given time is approximately 0.
This approach is not very accurate and can lead us to false negative conclusions (as in reality there will be a concentration gradient, with highest conentration on the cell wall).
<img src="http://www.openwetware.org/images/4/42/Outer_Volume.JPG" alt="Control volume (Volume of B.sub to be excluded. x indicates the distance travelled by AIPs from the surface in time t)">
Difussion distance was calculated using Fick's 1st Law: x=√2Dt, where: x - diffusion distance, D - diffusion constant, t - time of diffusion
Daverage = 10.7x10-11 m2s-1 for a protein in agarose gel for pH=5.6 <a href="http://www.sciencedirect.com/science?_ob=MImg&_imagekey=B6V5N-4B3MXDC-2-K&_cdi=5791&_user=217827&_pii=S1369703X03002377&_origin=search&_coverDate=07%2F01%2F2004&_sk=999809998&view=c&wchp=dGLzVtb-zSkzS&md5=c17d0e7320f03931006f9b1a10a438b9&ie=/sdarticle.pdf">[3]</a>
t = 8s (arbitrarily chosen)
This gives: x = 4.14x10-5m
The control volume can be calculated by adding 2x to the length and the diamter of the cell.
This gives a control volume (CV) = 4.81x10-7m3
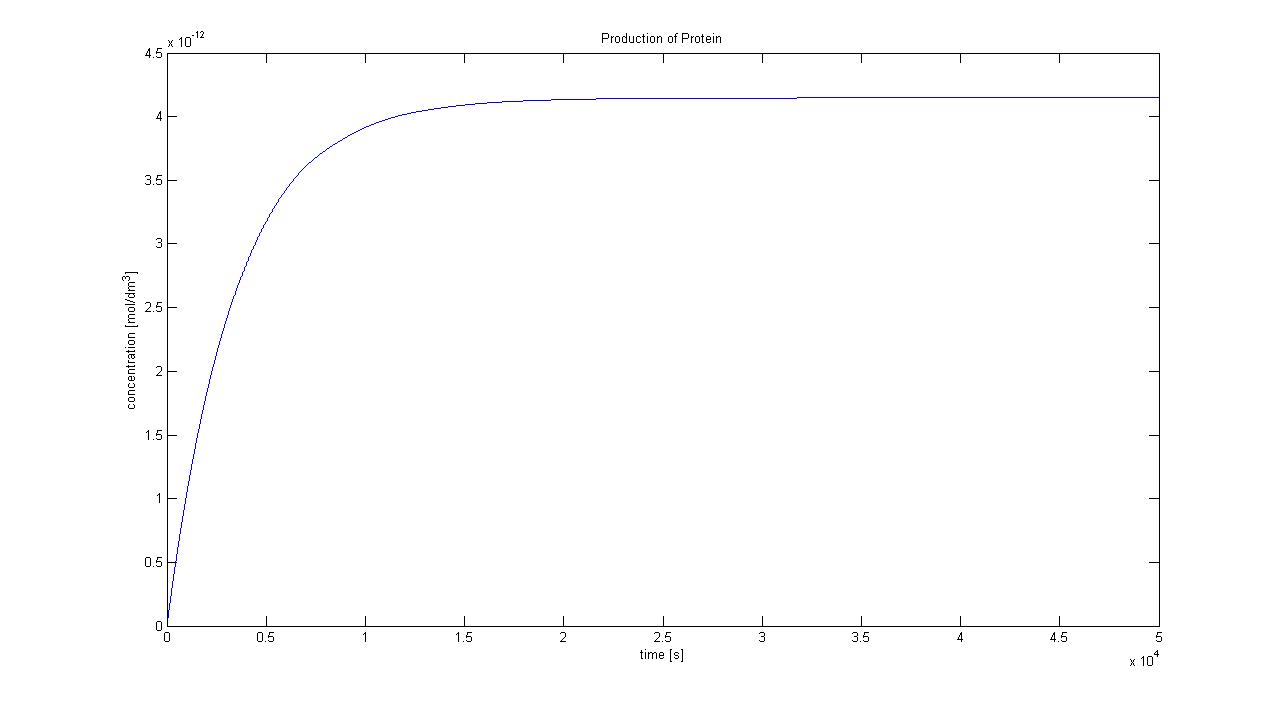
<img src="http://www.openwetware.org/images/7/78/Protein_in_control_volume.jpg" alt="Production of protein by transcription and translation in control volume">
Protein production in Control Volume (23/08/2010)
The previously determined constants of protein production in B.sub to obtain the concentration of proteins are not valid in the Control Volume. It has to be adjusted (multiplied) by the following factor:
factor=Vbacillus/VCV = 5.7974x10-6 (for the particular numbers presented above)
Control volume final choice (23/08/2010)
We realized that our initial choice of control volume was not accurate because this assumption was treating bacteria as the medium. However, in reality bacteria live in colonies very close to each other. They are much closer to each other than the diffusion distance (1.9596x10−5m) derived above even if placed in water solution.
Using CFU to estimate the spacing between cells (24/08/2010)
CFU stands for Colony-forming unit. It is a measure of bacterial numbers. For liquids, CFU is measured per ml.
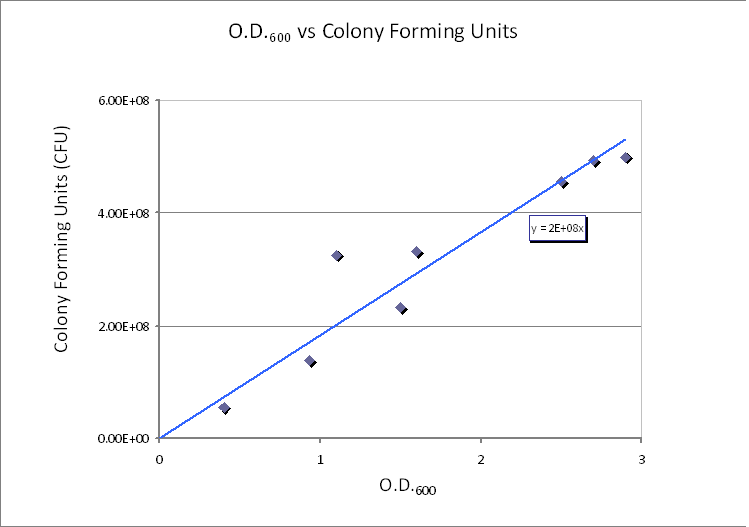
Since we already have data of CFU/ml from the Imperial iGEM 2008, this is an easy way to estimate the number of cells in a given volume using a spectrometer at 600nm wavelength.
The graph below is taken from the Imperial iGEM 2008 Wiki page <a hred="http://2008.igem.org/Imperial_College/18_September_2008">[4]</a>.
<img src="http://www.openwetware.org/images/b/bf/Calibration-Dilution.PNG" alt="CFU/ml vs. Optical Density at 600nm (OD600)">
The graph shows values of CFU/ml for different optical densities. The range of CFU/ml is therefore between 0.5x10^8 - 5x108.
In this calculation, we will assume that only one cell will grow and become one colony (i.e. no more than one cell will form no more than one colony). Therefore, the maximum number of cells in 1ml of solution is 5x108. Taking the volume of 1 ml = 10-3 dm3 and dividing by the (maximum) number of cells in 1ml gives the average control volume (CV) around each cell: 2x10-12 dm3/cell. For simplicity, we choose the control volume to be cubic. Taking the third root of this value gives the length of each side of the control volume. Side length of CV = y = 1.26x10-4 dm = 1.26x10-5 m.
Choice of Control Volume allows simplifications (24/08/2010)
- Firstly, assume that the cells will be placed in the centre of the CV. Hence, the protein (aftr cleavage) will have an average distance off y/2 to travel in order to cross the boundary of CV. This is calculated to happen within 0.18s. Even if the bacterium was not placed in the centre of the CV, the protein will travel from one end of the cube to the other in less than a second (~0.74s). Hence, it will take between 0.18 and 0.74s for the concentration of AIPs around the cell to be uniform. Noting that these time values are really small, we can approximate our model to have a uniform concentration across the volume. In this way we are underestimating the value of AIP concentration right next to the cell's surface. Hence, we are overestimating the time required for the AIP concentration to reach the threshold level.
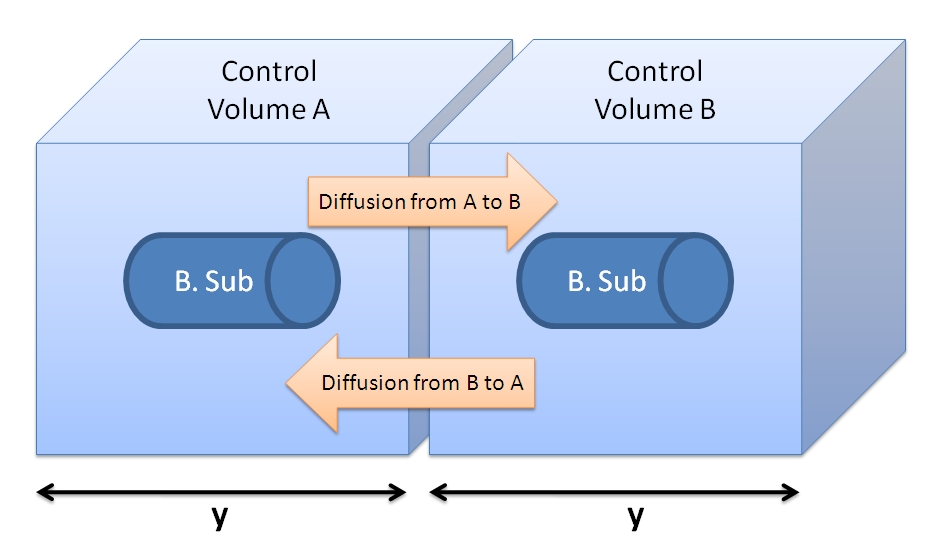
- We can neglect the diffusive fluxes across the CV border (see figure below). Assuming that adjacent cells are producing the peptide at the same rate and that the concentration of TEV is the same around the cell, then the fluxes should be of the same value giving a net flux of zero. Hence, we can neglect diffusion and have our model limited to one bacterium.
<img src="http://www.openwetware.org/images/f/f9/Control_Volume_and_fluxes.JPG" alt="Figure showing 2 cells with their control volumes.">
Conditions resulting from our assumptions
Most of our assumptions in choice of control volume were possible due to careful choice of cell density = 5x108 CFU/ml.
If the density is changed too much, then our simplifications might not hold any longer. However, this does not mean that our system cannot function for lower cell densities. Our model might not be very accurate for predicting situations with cell densities that are much higher or lower than 5x108 CFU/ml.
It was decided that the model should not be used for cell densities lower 107 CFU/ml. Below that value AIP takes more than a 1 second to diffuse accross half of a side-length of the control volume (assuming that the cell is inside control volume). We agreed that below 10^7 CFU/ml the approximation about uniform concentration throughout the control volume could be wrong and that the concentration gradients could become significant. If our model were applied to this particular situation, it would possibly overestimate the time taken to activate the receptor.
It is not possible to increase cell density by more than 109 CFU/ml</math> because of the size of a cell.
Matlab Simulation (24/08/2010)
<a href="http://www.openwetware.org/wiki/Image:Wiki_Model_Protein_Display.txt">Here is the Matlab code for the Matlab simulation</a>
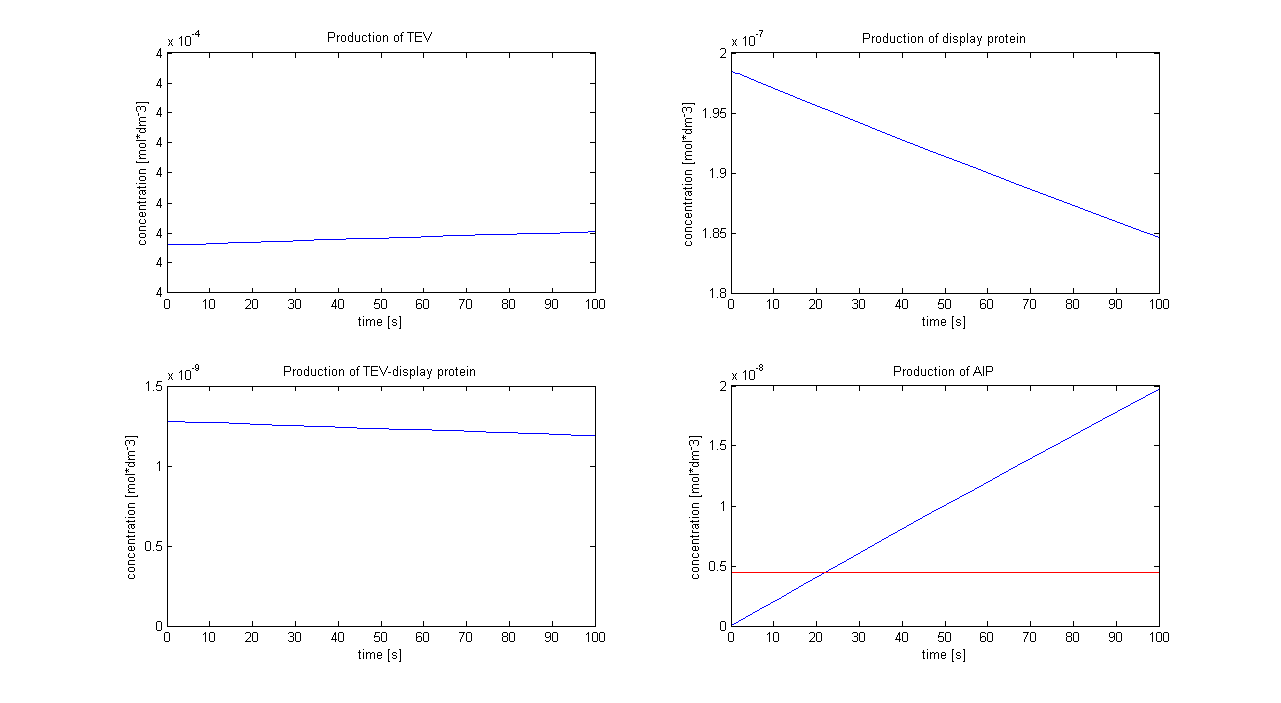
<img src="http://www.openwetware.org/images/7/77/Protein_display.png" alt="Graphs showing the simulation using [TEV]0 = 4x10-4 mol/dm3. The graph on the right hand-side below shows that the AIP threshold (red line) is reached after 22 s.">
Sensitivity of our model (24/08/2010)
- Changing initial concentration of TEV
- Changing the production rate
- Changing production rate
- Changing control volume
Whether the threshold concentration of AIP is reached is highly dependent on the initial concentration of TEV. The smallest initial concentration of TEV, [TEV>]<sub0, for which the threshold is reached is 6.0x10-6mol/dm3. On the grap below it can be seen that the optimal [TEV]0 is a concentration higher than 10-4mol/dm3, which corresponds to the threshold being reached within 1.5 minutes.
<img src="http://www.openwetware.org/images/d/df/AIP_Threshold_concentration.PNG" alt="Graph showing when threshold AIP concentration is reached (for different initial TEV concentrations). Notice log-log scale."/>
One order of magnitude change in the production rate results in at least 50s delay of the AIP concentration reaching the threshold concentration.
Changing the production rate influences the time duration of the AIP concentration above the threshold level. The higher it is, the shorter the receptor will be activated (at extreme values, AIP concentration does not reach the threshold). However, the production rate has not much influence on how fast the threshold will be reached.
Our model is extremely sensitive to this factor. One order of magnitude change in CV results in several orders of magnitude change in AIP concentration. Hence, special care should be taken in determination of this value. If the model is to be compared with the experimental results, the CFU/ml has to be the same as the one used in the model. Otherwise, the CV has to be readjusted.
Risk of False positives (31/08/2010)
It was pointed out that we should assess the risk of false positive activation of the receptor. We are particularly concerned about the display protein not binding to the cell wall, but instead diffusing into the extra-cellular environment.
In order to be able to assess the risk of false positives, we need to do further research into the affinity of AIP with attached linker and transmembrane proteins for the receptor as compared to the affinity of the AIP itself for the receptor.
This paper <a href="http://jb.asm.org/cgi/content/full/186/10/3078">[5]</a> might have some information on affinity comparison.
We need to know how proteins are being transported from intracellular to transmembrane space. Understanding this concept could give us an idea of what could go wrong.
References
- Havarstein, L., Coomaraswamy, G. & Morrison, D. (1995) An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proc. Natl. [Online] 92, 11140-11144. Available from: http://ukpmc.ac.uk/backend/ptpmcrender.cgi?accid=PMC40587&blobtype=pdf [Accessed 27th August 2010]
- Kobayashi, G. et al (2000) Accumulation of an artificial cell wall-binding lipase by Bacillus subtilis wprA and/or sigD mutants. FEMS Microbiology Letters. [Online] 188(2000), 165-169. Available from: http://onlinelibrary.wiley.com/doi/10.1111/j.1574-6968.2000.tb09188.x/pdf [Accessed 27th August 2010]
- Gutenwik, J., Nilsson, B. & Axelsson, A. (2003) Determination of protein diffusion coefficients in agarose gel with a diffusion cell. Biochemical Engineering Journal. [Online] 19(2004), 1-7. Available from: http://www.sciencedirect.com/science?_ob=MImg&_imagekey=B6V5N-4B3MXDC-2-K&_cdi=5791&_user=217827&_pii=S1369703X03002377&_origin=search&_coverDate=07%2F01%2F2004&_sk=999809998&view=c&wchp=dGLzVtb-zSkzS&md5=c17d0e7320f03931006f9b1a10a438b9&ie=/sdarticle.pdf [Accessed August 20th 2010]
- Imperial College London (2008) Biofabricator Subtilis - Designer Genes. [Online] Available from: http://2008.igem.org/Imperial_College/18_September_2008 [Accessed 1st September 2010]
- Knutsen, E., Ween, O. & Havarstein, L. (2003) Two Separate Quorum-Sensing Systems Upregulate Transcription of the Same ABC Transporter in Streptococcus pneumoniae. Journal of Bacteriology. [Online] 186(10), 3078-3085. Available from: http://jb.asm.org/cgi/reprint/186/10/3078 [Accessed 1st September 2010]
</body> </html>

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}